Author: Lynne Uhl, MD
Section Editor: Arthur J Silvergleid, MD
Literature review current through: Apr 2020. | This topic last updated: Sep 20, 2019.
Аллоиммунизация - это образование в материнском организме антител к антигенам плодовых эритроцитов (преимущественно к резус-антигену (D), реже к АГ Duffy и Kell) или тромбоцитам. Изосерологическая несовместимость (изоиммунизация) – это образование в крови матери антител в ответ на проникновение плодовых антигенов, несовместимых либо по системе АВО, либо по резус-фактору. Изоиммунизация по системе АВ0 развивается в том случае, если мать имеет 0(I) группу крови, а плод – А (II) или В (III) группу. Антитела к А- и В- антигенам всегда присутствуют в организме человека с первой группой крови, поэтому предварительной сенсибилизации для их образования не требуется.
С 38 дня беременности на циркулирующих эритроцитах плода появляется резус-антиген, но вероятность сенсибилизации в ранние сроки не превышает 5%. Помимо резус-антигена на эритроцитах экспрессируется большое количество антигеный структур, ряд из которых обладают сильной иммуногенностью (таблица).
Таблица. Антигенные структуры эритроцита


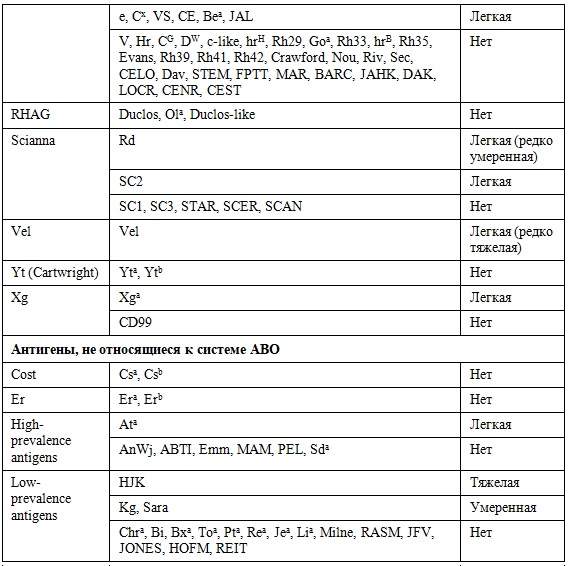
Примечание:
- Легкое течение ГБПиН: трансфузионная постнатальная терапия
- Умеренное течение ГБПиН: показано провдение заменного переливания крови
- Тяжелое течение ГБРиН: гидропс плода, требуется внутриутробное переливание отмытых эритроцитов.
Наиболее часто выявляются анти-D, анти- c и анти- Kell антитела. Реже аллоиммунизация развивается к эритроцитарным антигенам Е, Fy, jk, C и Се. Контакт с антигенными структурами запускает образование антител, относящихся к 4 классам IgG, но только 1 и 3 вызывают развитие тяжелых форма ГБПиН. IgG2 и IgG4 не играют значимой роди в развитии ГБП. Клинически значимая сенсибилизация развивается примерно у 1% женщин.
Система резус содержит 48 антигенов, наиболее часто встречающимися являются D, d, C, c, E, e и G. Кроме того, возможны различные вариации антигенов. Например, у резус-негативной женщины (подразумевается отсутствие D антигена) могут образовываться антитела к другим структурным антигенам системы резус, например анти-С, с, Е или е. В связи с тем, что у пациентки нет анти-D – ей будет проведена профилактическая иммунизация, но она не предупредит развитие аллоиммунизации к малым антигенным структурам эритроцитов плода.
При развитии анти-К аллоиммунизации, запускается не только внутриклеточный гемолиз, но и происходит подавление эритропоэза в костном мозге плода. Сложность заключается в том, что титр анти-К-антител не отражает степень тяжести болезни и риска развития тяжелых форм ГБП. После родов, гемолиз продолжается до момента исчезновения из кровотока материнских антител (максимальная длительность циркуляции АТ 12 недель). Гипербилирубинемия у плода не имеет большого значения, так как выведение избытков билирубина осуществляется с участием материнского организма. После родов гипербилирубинемия новорожденного является причиной развития ядерной желтухи с поражением структур головного мозга.
Lewis-аллоиммунизация не ассоциирована с развитием гемолитической болезни плода и новорожденного. Эта система включает Le(a), Le(b) и не является истинными антигенными структурами эритроцитов. Данные антигены экспрессируются другими тканями и циркулируют в связанном с эритроцитами состоянии.
Kell-аллоиммунизация встречается у 9% людей кавказской национальности и является причиной 10% тыжелых форм ГБП. Развитие анти-К-сенсибилизации чаще всего происходит при переливании компонентов крови без учета Kell-статуса. Частота анти- Kell алоиммунизации составляет 0.1-0.2%.
Duffy - антиген Fy(a) и Fy(b) может присутствовать в различных вариантах: Fy(a+b+), Fy(a-b-), Fy(a+b-), or Fy(a-b+), но только анти-Fy(a) антитела ассоциированы с ГБП.
MNS - система содержит M, N, S, s и U антигены. Умеренное или легкое течение ГБП возможно при аллоиммунизации к -S, -s и –U антигенам.
P -система содержит P1 и P2 антигены, которые встречаются у 79 и 21% кавказцев соответственно. При наличии у женщины анти -P1+P+P(k) антител значительно повышен риск ГБП и привычного невынашивания беременности.
Уровень анти-D антител ниже 4 МЕ/мл (титр 1:4) не свидетельствует о риске развития гемолитической болезни плода. При уровне 4-15 МЕ/мл – риск умеренно повышен, а при уровне антител более 15 МЕ/мл – значительно повышен.
Анти-с антитела относятся к антителам системы резус, но риск развития ГБП значительно ниже, чем при наличии анти-D сенсибилизации. При уровне антител до 7.5 МЕ/мл – необходимо продолжить наблюдение. При уровне 7.5-20 МЕ/мл – риск ГБП повышен умеренно, проводится оценка скоростей кровотока в СМА. При уровне выше 20 МЕ/мл – риск ГБП повышен значительно и пациентка должна быть направлена в перинатальный медицинский центр для принятия решения о проведении кордоцентеза и внутриутробного переливания отмытых эритроцитов.
Анти-Е антитела свидетельствуют о риске развития гемолитической анемии у плода только при их титре выше 1:32.
Литература:
1. Daniels G, Reid ME. Blood groups: the past 50 years. Transfusion 2010; 50:281.
2. Flegel WA. Pathogenesis and mechanisms of antibody-mediated hemolysis. Transfusion 2015; 55 Suppl 2:S47.
3. Technical Manual, 19th edition, Fung M, Eder AF, Spitalnik SL, et al (Eds), AABB Press, Bethesda, MD 2017.
4. Denomme GA. The structure and function of the molecules that carry human red blood cell and platelet antigens. Transfus Med Rev 2004; 18:203.
5. Curtis BR, Edwards JT, Hessner MJ, et al. Blood group A and B antigens are strongly expressed on platelets of some individuals. Blood 2000; 96:1574.
6. Curtis BR, Fick A, Lochowicz AJ, et al. Neonatal alloimmune thrombocytopenia associated with maternal-fetal incompatibility for blood group B. Transfusion 2008; 48:358.
7. Kamphuisen PW, Eikenboom JC, Bertina RM. Elevated factor VIII levels and the risk of thrombosis. Arterioscler Thromb Vasc Biol 2001; 21:731.
8. Morange PE, Tregouet DA, Frere C, et al. Biological and genetic factors influencing plasma factor VIII levels in a healthy family population: results from the Stanislas cohort. Br J Haematol 2005; 128:91.
9. Trégouët DA, Heath S, Saut N, et al. Common susceptibility alleles are unlikely to contribute as strongly as the FV and ABO loci to VTE risk: results from a GWAS approach. Blood 2009; 113:5298.
10. Reid ME, Bird GW. Associations between human red cell blood group antigens and disease. Transfus Med Rev 1990; 4:47.
11. Wolpin BM, Chan AT, Hartge P, et al. ABO blood group and the risk of pancreatic cancer. J Natl Cancer Inst 2009; 101:424.
12. Amundadottir L, Kraft P, Stolzenberg-Solomon RZ, et al. Genome-wide association study identifies variants in the ABO locus associated with susceptibility to pancreatic cancer. Nat Genet 2009; 41:986.
13. Risch HA, Yu H, Lu L, Kidd MS. ABO blood group, Helicobacter pylori seropositivity, and risk of pancreatic cancer: a case-control study. J Natl Cancer Inst 2010; 102:502.
14. Dipta TF, Hossain AZ. The Bombay blood group: are we out of risk? Mymensingh Med J 2011; 20:536.
15. Human Blood Groups, 3, Geoff Daniels (Ed), Wiley-Blackwell, 2013.
16. Roath S, Todd CE, Shaw D. Transient acquired blood group B antigen associated with diverticular bowel disease. Acta Haematol 1987; 77:188.
17. Daniel-Johnson J, Leitman S, Klein H, et al. Probiotic-associated high-titer anti-B in a group A platelet donor as a cause of severe hemolytic transfusion reactions. Transfusion 2009; 49:1845.
18. Julius CJ, Wade M, Waheed A, et al. Common variable immunodeficiency: diagnosis by absent ABO reverse type. Immunohematology 1997; 13:80.
19. Linz WJ, Currie RT. ABO discrepancy: reverse type. Transfusion 2007; 47:1.
20. Beattie KM. Perspectives on some usual and unusual ABO phenotypes. In: A seminar on antigens on blood cells and body fluids, Bell CA (Ed), American Association of Blood Banks, Washington, DC 1980. p.97.
21. The ABO Blood Group System. In: Modern Blood Banking and Transfusion Practices, Harmening DM (Ed), 1999. p.113.
22. Josephson CD, Castillejo MI, Grima K, Hillyer CD. ABO-mismatched platelet transfusions: strategies to mitigate patient exposure to naturally occurring hemolytic antibodies. Transfus Apher Sci 2010; 42:83.
23. Van Kim CL, Colin Y, Cartron JP. Rh proteins: key structural and functional components of the red cell membrane. Blood Rev 2006; 20:93.
24. Avent ND, Reid ME. The Rh blood group system: a review. Blood 2000; 95:375.
25. Westhoff CM. The Rh blood group system in review: a new face for the next decade. Transfusion 2004; 44:1663.
26. Reid MR, Lomas-Francis C. The blood group antigen facts book, Harcourt Brace and Company, 1997.
27. Denomme GA, Wagner FF, Fernandes BJ, et al. Partial D, weak D types, and novel RHD alleles among 33,864 multiethnic patients: implications for anti-D alloimmunization and prevention. Transfusion 2005; 45:1554.
28. Kumpel B. Are weak D RBCs really immunogenic? Transfusion 2006; 46:1061.
29. Shao CP. Transfusion of RhD-positive blood in "Asia type" DEL recipients. N Engl J Med 2010; 362:472.
30. Chou ST, Jackson T, Vege S, et al. High prevalence of red blood cell alloimmunization in sickle cell disease despite transfusion from Rh-matched minority donors. Blood 2013; 122:1062.
31. Karafin MS, Field JJ, Gottschall JL, Denomme GA. Barriers to using molecularly typed minority red blood cell donors in support of chronically transfused adult patients with sickle cell disease. Transfusion 2015; 55:1399.
32. Muller CL, Schucker JL, Boctor FN. When anti-G and anti-C antibodies masquerade as anti-D antibody. J Matern Fetal Neonatal Med 2011; 24:193.
33. O'Brien KL, Haspel RL, Uhl L. Anti-D alloimmunization after D-incompatible platelet transfusions: a 14-year single-institution retrospective review. Transfusion 2014; 54:650.
34. Borén T, Falk P, Roth KA, et al. Attachment of Helicobacter pylori to human gastric epithelium mediated by blood group antigens. Science 1993; 262:1892.
35. Ilver D, Arnqvist A, Ogren J, et al. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science 1998; 279:373.
36. Hellberg A, Westman JS, Olsson ML. An update on the GLOB blood group system and collection. Immunohematology 2013; 29:19.
37. Yu LC, Twu YC, Chou ML, et al. The molecular genetics of the human I locus and molecular background explain the partial association of the adult i phenotype with congenital cataracts. Blood 2003; 101:2081.
38. Hossaini AA. Neutralization of Lewis antibodies in vivo and transfusion of Lewis incompatible blood. Am J Clin Pathol 1972; 57:489.
39. WORLLEDGE SM, ROUSSO C. STUDIES ON THE SEROLOGY OF PAROXYSMAL COLD HAEMOGLOBINURIA (P.G.H.), WITH SPECIAL REFERENCE TO ITS RELATIONSHIP WITH THE P BLOOD GROUP SYSTEM. Vox Sang 1965; 10:293.
40. Reid ME. MNS blood group system: a review. Immunohematology 2009; 25:95.
41. Wikman A, Edner A, Gryfelt G, et al. Fetal hemolytic anemia and intrauterine death caused by anti-M immunization. Transfusion 2007; 47:911.
42. Bakhtary S, Gikas A, Glader B, Andrews J. Anti-Mur as the most likely cause of mild hemolytic disease of the newborn. Transfusion 2016; 56:1182.
43. Symmans WA, Shepherd CS, Marsh WL, et al. Hereditary acanthocytosis associated with the McLeod phenotype of the Kell blood group system. Br J Haematol 1979; 42:575.
44. Marsh WL, Marsh NJ, Moore A, et al. Elevated serum creatine phosphokinase in subjects with McLeod syndrome. Vox Sang 1981; 40:403.
45. Neote K, Mak JY, Kolakowski LF Jr, Schall TJ. Functional and biochemical analysis of the cloned Duffy antigen: identity with the red blood cell chemokine receptor. Blood 1994; 84:44.
46. Afenyi-Annan A, Kail M, Combs MR, et al. Lack of Duffy antigen expression is associated with organ damage in patients with sickle cell disease. Transfusion 2008; 48:917.
47. Miller LH, Mason SJ, Clyde DF, McGinniss MH. The resistance factor to Plasmodium vivax in blacks. The Duffy-blood-group genotype, FyFy. N Engl J Med 1976; 295:302.
48. McDougall DC, McGregor M. Jk:-3 red cells have a defect in urea transport: a new urea-dependent lysis test. Transfusion 1988; 28:197.
49. Wester ES, Johnson ST, Copeland T, et al. Erythroid urea transporter deficiency due to novel JKnull alleles. Transfusion 2008; 48:365.
50. Heaton DC, McLoughlin K. Jk(a-b-) red blood cells resist urea lysis. Transfusion 1982; 22:70.
51. Sands JM, Gargus JJ, Fröhlich O, et al. Urinary concentrating ability in patients with Jk(a-b-) blood type who lack carrier-mediated urea transport. J Am Soc Nephrol 1992; 2:1689.
52. Parsons SF, Lee G, Spring FA, et al. Lutheran blood group glycoprotein and its newly characterized mouse homologue specifically bind alpha5 chain-containing human laminin with high affinity. Blood 2001; 97:312.
53. An X, Gauthier E, Zhang X, et al. Adhesive activity of Lu glycoproteins is regulated by interaction with spectrin. Blood 2008; 112:5212.
54. Rowe PC, McLean RH, Wood RA, et al. Association of homozygous C4B deficiency with bacterial meningitis. J Infect Dis 1989; 160:448.
55. Dunckley H, Gatenby PA, Hawkins B, et al. Deficiency of C4A is a genetic determinant of systemic lupus erythematosus in three ethnic groups. J Immunogenet 1987; 14:209.
56. Pasvol G, Anstee D, Tanner MJ. Glycophorin C and the invasion of red cells by Plasmodium falciparum. Lancet 1984; 1:907.
57. Reynolds MV, Vengelen-Tyler V, Morel PA. Autoimmune hemolytic anemia associated with autoanti-Ge. Vox Sang 1981; 41:61.
58. Mochizuki T, Tauxe WN, Ramsey G. In vivo cross-match by chromium-51 urinary excretion from labeled erythrocytes: a case of anti-Gerbich. J Nucl Med 1990; 31:2042.
59. Hildebrandt M, Hell A, Etzel F, et al. Determination and Successful Transfusion of Anti-Gerbich-Positive Red Blood Cells in a Patient with a Strongly Reactive Anti-Gerbich Antibody. Infusionsther Transfusionsmed 2000; 27:154.
60. Noumsi GT, Tounkara A, Diallo H, et al. Knops blood group polymorphism and susceptibility to Mycobacterium tuberculosis infection. Transfusion 2011; 51:2462.












